Bảo tồn, lưu giữ nguồn gen Vi sinh vật thú y
STNN - Nguồn gen vi sinh vật thú y là bộ phận không thể thiếu đối với công tác bảo vệ sức khỏe cộng đồng thông qua bảo vệ sức khỏe vật nuôi.
STNN - Nguồn gen vi sinh vật thú y (VSVTY) là bộ phận không thể thiếu đối với công tác bảo vệ sức khỏe cộng đồng thông qua bảo vệ sức khỏe vật nuôi. Đây là công cụ, nguyên liệu cho việc chế tạo các chế phẩm sinh học (bao gồm vắc xin, kháng huyết thanh, kháng nguyên và các chế phẩm dùng cho chẩn đoán khác...).
 Từ trên ba mươi năm nay, Trung tâm Kiểm nghiệm thuốc Thú y TW1 được giao nhiệm vụ bảo tồn, lưu giữ nguồn gen VSVTY. Đây là nhiệm vụ cấp thiết và thường xuyên của Trung tâm nhằm phục vụ công tác đánh giá chất lượng vắc xin và các chế phẩm sinh học trong nước, nhập khẩu. Bên cạnh đó, Trung tâm còn cung cấp một số chủng giống vi sinh vật cho chẩn đoán, nghiên cứu và giảng dạy khi có sự đồng ý của cơ quan có thẩm quyền.
Từ trên ba mươi năm nay, Trung tâm Kiểm nghiệm thuốc Thú y TW1 được giao nhiệm vụ bảo tồn, lưu giữ nguồn gen VSVTY. Đây là nhiệm vụ cấp thiết và thường xuyên của Trung tâm nhằm phục vụ công tác đánh giá chất lượng vắc xin và các chế phẩm sinh học trong nước, nhập khẩu. Bên cạnh đó, Trung tâm còn cung cấp một số chủng giống vi sinh vật cho chẩn đoán, nghiên cứu và giảng dạy khi có sự đồng ý của cơ quan có thẩm quyền.
Xuất phát từ thực tiễn trên, TS. Nguyễn Trung Tiến và các cộng sự tại Trung tâm kiểm nghiệm thuốc thú y TW1 - Cục Thú y thuộc Bộ Nông nghiệp và Phát triển nông thôn đã thực hiện nghiên cứu “Bảo tồn, lưu giữ nguồn gen Vi sinh vật thú y” với mục tiêu: Bảo tồn và lưu giữ nguồn gen, góp phần sử dụng hiệu quả, bền vững nguồn gen vi sinh vật thú y và tiến tới xây dựng ngân hàng gen; Khai thác và sử dụng hiệu quả phục vụ công tác bảo vệ sức khỏe đàn gia súc, gia cầm; Tăng khả năng cạnh tranh trong xu thế hội nhập quốc tế.
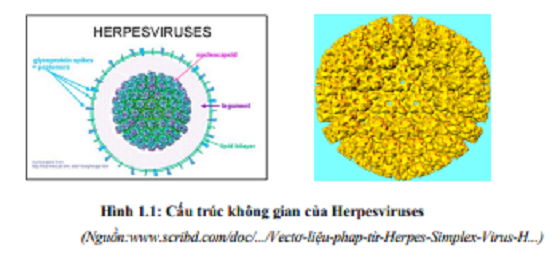
Về hình thái và cấu tạo tất cả mọi Herpesvirus đều giống nhau, đều có lớp vỏ bọc ngoài cùng là các nucleocapsid, theo P. Spradbrow (1997) rất mẫn cảm với ether và chloroform. Herpesvirus là loại virus ADN, có hệ gen là ADN sợi đôi có độ dài khoảng 150-170 kb giống như nhiều loại virusthuộc họ Herpesviridae khác. Virus dịch tả vịt có cấu trúc capsid đối xứng khối 20 mặt với 162 capsomere khối lượng phân tử là 119x106 Dalton, phân bố thành phần nucleotide trong hệ gen thiên về G+C (Guanine và Cytosine), chiêm tỷ lệ 64,3%, cao hơn rất nhiều so với các Herpesvirus khác của gia cầm, ví dụ như virusgây bệnh Marek chẳng hạn, hay virus gây bệnh viêm thanh khí quản truyền nhiễm (Gardner và cs, 1993).
Khi quan sát tế bào nhiễm virus dịch tả vịt sau 48h bằng kính hiển vi điện tử, Breese và Dardiri (1968) đã thấy rằng có 2 loại hình thái virus dịch tả vịt. Ở trong nhân tế bào chủ chúng có đường kính khoảng 91nm với nhân khoảng 48nm. Ở trong nguyên sinh chất chúng có đường kính khoảng 181nm với nhân khoảng 74nm.
Bệnh viêm túi Fabricius truyền nhiễm là do một loại virus có tên gọi là virus Gumboro gây lên. Virus Gumboro rất nhỏ bé và không có vỏ bọc bên ngoài cùng. Cấu trúc đơn giản nhưng hoàn thiện vì thế chúng có khả năng chống chịu lại sự tác động bất lợi của môi trường. Mặt khác, do kích thước nhỏ bé và cấu trúc linh hoạt (hệ gen đơn giản), virus thích ứng xâm nhập và truyền lây trong quần thể vật chủ một cách nhanh chóng và dễ dàng thoát khỏi miễn dịch tự nhiên của cơ thể.
Về mặt cấu trúc, virus Gumboro có đối xứng khối bao gồm 32 capxome tạo nên vỏ capxit và có đường kính 58- 60 nm. Capxit của virus là lớp vỏ protein đơn lớp. Virus không có vỏ bọc ngoài cùng, mà thông thường lớp vỏ đó có cấu trúc từ lipoprotein rất mẫm cảm với ether và chloroform.
Virus Gumboro bao gồm một hệ gen chứa hai sợi ARN và phân bố trong hai phân đoạn khác nhau (Kibenge và cộng sự, 1988). Hai phân đoạn trong hệ gen ARN của virus có tên gọi là phân đoạn A và B. Phân đoạn B có độ dài khoảng 2800 nucleotit, chỉ chứa duy nhất một cấu trúc gen mã hóa cho sự tổng hợp một protein duy nhất có tên gọi là VP1 (VP = viral protein). VP1 là protein có hoạt tính sinh học chịu trách nhiệm là enzym ARN- polymerase của virus. Enzym này có vai trò xúc tác trong quá trình tổng hợp nguyên liệu ARN, vật liệu di truyền của virus.
Phân đoạn A trong hệ gen của virus Gumboro có độ dài tổng cộng là 3400 nucleotit bao gồm hai bộ phận gen tổng hợp. Bộ phận thứ nhất là mộ cấu trúc đơn gen (monocistronic) mã hóa cho một tiền protein có phân tử lượng là 110 kDa mà trong quá trình tiếp theo sẽ được phân cắt thành các protein cấu trúc tên gọi là VP2, VP3, VP4 (Moller và Becht, 1982; Azad và cộng sự, 1985; 1987; Hudson và cộng sự, 1986; Kibenge và cs. 1997). Bộ phận thứ hai là một phần ADN trong phân đoạn A, mã hóa cho một loại protein khác có tên là VP5, mà chuỗi ADN này lồng vào trong phần gen của VP2. Do vậy hai loại protein này sử dụng chung một phần chuỗi nucleotit làm thành phần của mình nhưng bộ ba mã hóa cho thành phần axit amin là khác nhau. VP5 là một protein có trọng lượng phân tử bé (17kDa) mới được phát hiện gần đây có chức năng trong quá trình điều hòa sao chép và tổng hợp protein (Mundt và cs, 1995).
VP2 và VP3 là protein cấu trúc, cấu tạo nên thành phần ngoài cùng của virus - đó là capxit. Trong cấu trúc capxit, VP2 trình diện lên bề mặt, là thành phần protein bề mặt, còn VP3 lặn sâu vào trong, là thành phần cấu tạo bên trong. VP4 chính là proteaza, một loại enzym có chức năng phân cắt protein- có vai trò trong cắt rời chuỗi polypeptit do toàn bộ phân đoạn A tổng hợp gọi là protein chung (polyprotein). VP4 được tổng hợp do toàn bộ cấu trúc đa gen (polycistronic) của phân đoạn A (Azad và cs, 1987; Kibenge và cs, 1997; Granzan và cs, 1997). VP2 đã được chứng minh là một loại protein có tính kháng nguyên và có chất bảo vệ virus. Do vậy, VP2 đại diện cho tính động lực và tính gây bệnh của virus. VP2 bao gồm một vùng được gọi là vùng kháng nguyên, có vai trò trong quá trình kích thích cơ thể động vật bị nhiễm bệnh sản sinh ra kháng thể trung hòa (Fahey và cs, 1989).
Sự biến đổi bất kỳ một axit amin nào trong VP2 đều có thể dẫn đến sự thay đổi nhất định của tính kháng nguyên và tính gây bệnh. Tuy nhiên, có một vùng hẹp trong cấu trúc gen của VP2-gọi là vùng “siêu biến đổi” (hypervariable region) – có thành phần nucleotit và axit amin thay đổi trong các chủng khác nhau, định vị trí từ các nucleotit và axit amin thay đổi trong các chủng khác nhau, định vị trí từ các nucleotit thứ 703 đến nucleotit thứ 1176, bao gồm 474 nucleotit chịu trách nhiệm đặc biệt về tính kháng nguyên. Protein VP2 sử dụng vùng “quyết định kháng nguyên” này để kích thích cơ thể sản sinh kháng thể có chất lượng thuộc loại kháng thể tham gia phản ứng trung hòa VN (virus neutralization). Một dãy gồm 12-20 axit amin trong vùng “siêu biến đổi” gọi là epitope có cấu trúc vòm đặc biệt chịu trách nhiệm kích thích và kết hợp với kháng thể tương ứng. Nếu vùng “siêu biến đổi” có thành phần nucleotit biến đổi trong giới hạn nhất định, không hoặc rất ít làm thay đổi axit amin của vùng “siêu biến đổi” nói chung và dãy epitope nói riêng, thì tính miễn dịch vẫn được đảm bảo trong quan hệ tương tác kháng nguyên- kháng thể giữa các virus Gumboro tương ứng. Như vậy, VP2 chính là loại hình kháng nguyên ts (type-specific).
Sau thời gian nghiên cứu, đề tài đã thu được những kết quả như sau:
- Lưu giữ và bảo quản thường xuyên 42 nguồn gen vi sinh vật thú y.
- Đã kiểm tra, đánh giá định kỳ 6 giống VSV bao gồm: Tụ huyết trùng trâu bò (Pbu.1, Pb.2); Phó thương hàn lợn (Sc.4, Sc.5); Dịch tả vịt cường độc và Gumboro cường độc CVL52/70.
- Đã xác định sự có mặt của 2 gen Stn và InvA và giải mã gen InvA (1600 bp) của 2 giống vi khuẩn Phó thương hàn lợn (Sc.4, Sc.5).
- Đã tư liệu hoá - lập hồ sơ chi tiết được 06 giống VSV (Tụ huyết trùng trâu bò (Pbu.1, Pb.2); Phó thương hàn lợn (Sc.4, Sc.5); Dịch tả vịt cường độc và Gumboro cường độc CVL52/70).
- Đã sử dụng một số giống trong kiểm nghiệm, khảo nghiệm vắc xin và chế phẩm sinh học dùng trong thú y.
Có thể tìm đọc toàn văn Báo cáo kết quả nghiên cứu (mã số 18872/2021) tại Cục Thông tin khoa học công nghệ quốc gia.
Nguồn: vista.gov.vn
Link nội dung: https://sinhthainongnghiep.net.vn/bao-ton-luu-giu-nguon-gen-vi-sinh-vat-thu-y-a26307.html